The Shadows of the Mind |
by Roger Penrose |
If we are to believe that neurons are the only things that
control the sophisticated actions of animals,then the humble paramecium presents
us with a profound problem. For she swims about her pond with her numerous
hairlike legs - the cilia - darting in the direction of bacterial
food which she senses using a variety of mechanisms,or retreating at
the prospect of danger,ready to swim off in another direction. She can also
negotiate obstructions by swimming around them. Moreover,she can apparently
even learn from her past experiences - though this most remarkable
of her apparent faculties has been disputed by some. How is this all achieved
by an animal without a single neuron or synapse? Indeed,being but a single
cell,and not being a neuron herself,she has no place to accommodate such
accessories.
|
| A paramecium. Note the hair-like cilia that are used for swimming. These form the external extremities of the paramecium's cytoskeleton. |
Yet there must indeed be a complicated control system governing
the behaviour of a paramecium - or indeed other one-celled animals like amoebas
- but it is not a nervous system.The structure
responsible is apparently part of what is referred to as the
cytoskeleton. As it name
suggests,the cytoskeleton provides the framework that holds the cell in shape,but
it does much more.The cilia themselves are endings of the cytoskeletal fibres,but
the cytoskeleton seems also to contain the control system for the cell,in
addition to providing "conveyor belts" for the transporting of various molecules
from one place to another.In short,the cytoskeleton appears to play a role
for the single cell rather like a combination of skeleton,muscle
system,legs,blood circulatory system,and nervous system all rolled into
one!
It is the cytoskeleton's role as the cells "nervous system"
that will have the main importance for us here.For our own neurons are themselves
single cells,and each neuron has its own cytoskeleton! Does this mean
that there is a sense in which each individual neuron might itself have something
akin to its own "personal nervous system"? This is an intriguing issue,and
a number of scientists have been coming round to the view that something
of this general nature might actually be true.(See Stuart Hameroff's pioneering
1987 book Ultimate Computing: Biomolecular Consciousness and NanoTechnology;also
Hameroff and Wyatt (1982) and numerous articles in the new journal
Nanobiology.)
In order to address such issues,we should first glimpse the
basic organisation of the cytoskeleton.It consists of protein-like molecules
arranged in various types of structure:actin,
microtubules, and intermediate filaments.It is the
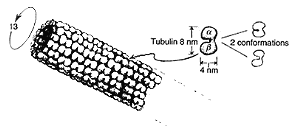
microtubules that will be our main concern here.They consist of hollow
cylindrical tubes,some 25nm in diameter on the outside and 14 nm on the
inside (where "nm"= "nanometre", which is 10-9 m),sometimes
organized into larger tubelike fibres that consist of nine doublets,triplets,or
partial triplets,of
microtubules,
organized in an arrangement with a fanlike cross-section,as indicated below,with
sometimes a pair of microtubules running down the centre.The paramecium's
cilia are structures of this kind.Each microtubule is itself a protein
polymer consisting of subunits referred to as tubulin.
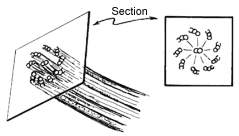 |
| Important parts of the cytoskeleton consist of bundles of tiny tubes (microtubules) organized in a structure with a fan-like cross-section. The paramecium's cilia are bundles of this nature. |
Each tubulin subunit is a "dimer" ie it consists of two
essentially separate parts called
a - tubulin and
b - tubulin each being
composed of about 450 amino acids. It is a globular protein pair,somewhat
"peanut shaped" and organized in a slightly skew hexagonal lattice along
the entirety of the tube,as indicated below. There are generally 13 columns
of tubulin dimers to each microtubule.Each dimer is about 8nm x 4nm x 4nm
and its atomic number is about 11 x 104 (which means that it has
about that many nucleons in it,so its mass,in absolute units,is about
10-14).
Each tubulin dimer,as a whole,can exist in (at least) two different
geometrical configurations - called conformations.In one of these,they bend
to about 30o to the direction of the microtubule. There is evidence
that these two conformations correspond to two different states of the dimer's
electric polarization,where these come about because an electron,centrally
placed at the a - tubulin
/b - tubulin juncture,
can shift from one position to the other.
The "control centre" of the cytoskeleton (if indeed this is
really an appropriate term) is a structure known as the centriole.This seems
to consist essentially of two cylinders of nine triplets of microtubules,where
the cylinders form a kind of separated "T"(See below right).
|
|
| A microtubule. It is a hollow tube,normally consisting of 13 columns of tubulin dimers. Each tubulin molecule is capable of (at least) two conformations. | The centriole (which appears to be the "control centre" of the cytoskeleton-if such exists) consists essentially of a separated "T" built from two bundles of microtubules. |
(The cylinders are similar,in a general way,to those that occur
in cilia).The centriole forms the critical part of a structure called the
microtubules organizing centre or centrosome.Whatever the role
of the centriole might be during the normal course of an ordinary cell's
existence,it has at least one fundamentally important task. At a critical
stage,each of the two cylinders in the centriole grows another,so as to make
two centriole "T"s that then separate from each other,each apparently
dragging a bundle of microtubules with it - although it would be more accurate
to say that each becomes a focal point around which microtubules assemble.
These microtubule fibres somehow connect the centriole to the separate
DNA strands in the nucleus (at central points,known
as their centromeres) and the DNA strands separate - initiate the extraordinary
process technically known as mitosis, which simply means cell division (see
below).
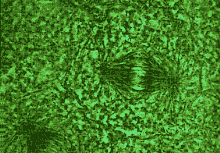 |
| In mitosis (cell division) the chromosomes separate, being pulled apart by microtubules. |
It may seem odd that there should be two quite different
"headquarters" in a single cell.On the one hand there is the
nucleus,where the fundamental genetic material of the cell resides,which
controls the cell's heredity and its own particular identity,and governs
the production of the protein materials of which the cell itself is composed.
On the other hand,there is the centrosome with its chief component
the centriole,which seems to be the focal point of the cytoskeleton,a
structure that apparently controls the cell's movements and its detailed
organization. The presence of these two different structures in eukaryotic
cells (the cells of all animals and almost all plants on this planet - but
excluding bacteria,blue-green algae,and viruses) is believed to be the result
of an ancient "infection" that took place some thousands of millions of years
ago. The cells that previously inhabited the earth were prokarytoic cells
that still exist today as bacteria and blue-green algae,and which possess
no cytoskeleton. One suggestion (Sagan 1976) is that some early prokaryotes
became entangled with - or,perhaps,"infected by" - some kind of spirochete,an
organism that swam with a whiplike tail composed of cytoskeleton proteins.
These mutually alien organisms subsequently grew to live permanently together
in a symbiotic relationship as single eukaryotic cells.Thus, these
"spirochetes" ultimately became cells' cytoskeletons - with al the implications
for the future evolution that thereby made us possible!
The organization of mammalian microtubules is interesting from
a mathematical point of view. The number 13 might seem to have no particular
mathematical significance,but this is not entirely so.It is one of the
famous
Fibonacci numbers:
0,1,1,2,3,5,8,13,21,34,55,89,144,.....
A sunflower head. As with many other plants,Fibonacci numbers feature strongly. In the outer regions,there are 89 clockwise and 55 anticlockwise spirals. Nearer the center we can find other Fibonacci numbers. |
|
where each successive number is obtained as a sum of the previous
two. This might be fortuitous, but Fibonacci
numbers are well known to occur frequently (at a much larger scale) in
biological systems. For example, in fir cones,sunflower heads,and palm tree
trunks,one finds spiral or helical arrangements involving the interpenetration
of right-handed and left-handed twists,where the number of rows for one
handedness and the number for the other handedness are two successive
Fibonacci numbers (see below).(As one examines the structures from one end
to the other,one may find that a "shunt" takes place,and the numbers then
shift to an adjacent pair of successive Fibonacci numbers.) Curiously,the
skew hexagonal pattern of microtubules exhibits a very similar feature -
generally of an even more precise organization - and it is apparently found
(at least normally) that this pattern is made up of 5 right-handed and 8
left-handed helical arrangements (see below).
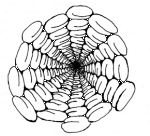
|
View down a microtubule! The 5 + 8 spiral arrangement
of the tubulins in this microtubule can be seen. |
|
Imagine a microtubule slit along its length, and then opened
out flat into a strip. We find that the tubulins are ordered in sloping lines
which rejoin at the opposite edge 5 or 8 places displaced (depending upon
whether the lines slope to the right or to the left).
|
In the diagram I have tried to indicate how this structure
might appear as actually "viewed" from within a microtubule. The number 13
features here in its role as the sum: 5+8. It is curious,also,that the double
microtubules that frequently occur seem normally to have a total of 21 columns
of tubulin dimers forming the outside boundary of the composite tube - the
next Fibonacci number! (However, one should not get carried away with such
considerations; for example,the "9" that occurs in the bundles of microtubules
in cilia and centrioles is not a Fibonacci number.) [But it is
the square of a Fibonacci number,but if one included all indices of Fibonacci's
a great many numbers would be involved increasing the likelihood of some
biological entity to have made use of it -LB]
Why do Fibonacci numbers arise in microtubule structure? In
the case of fir cones and sunflower heads,etc.,there are various plausible
theories - and Alan
Turing himself was someone who thought seriously about the subject
(Hodges 1983, p437).But it may well be that these theories are not appropriate
for microtubules, and different ideas are probably relevant at this level.
Koruga (1974) has suggested that these Fibonacci numbers may provide advantages
for the microtubule in its capacity as an "information processor".Indeed,Hameroff
and his colleagues have argued,for more than a decade,that microtubules may
play roles as cellular automata,where complicated
signals could be transmitted and processed along the tubes as waves of differing
electric polarization states of the tubulins. Recall that tubulin dimers
can exist in (at least) two different conformational states that can switch
from one to the other,apparently because of alternative possibilities for
their electric polarizations. The state of each dimer would be influenced
by the polarization states of each of its six neighbours (because of van
der Waals interactions between them) giving rise to certain specific rules
governing the conformation of each dimer in terms of the conformations of
its neighbours. This would allow all kinds of messages to be propagated and
processed along the length of each microtubule. These propagating signals
appear to be relevant to the way that microtubules transport various molecules
alongside them,and to the various interconnections between neighbouring
microtubules - in the from of bridge-like connecting proteins referred to
as MAPs (microtubule associated proteins) (see below).Koruga argues for a
special efficiency in the case of Fibonacci-number-related structure of the
kind that is actually observed for microtubules.There must indeed be some
good reason for this kind of organization in microtubules,since although
there is some variation in the numbers that apply to eukaryotic cells
generally,13 columns seems to be almost universal amongst mammalian
microtubules.
 |
| Microtubules tend to be interconnected with neighbouring ones by bridges of microtubule associated proteins (MAPs). |
What is the significance of microtubules for neurons? Each
individual neuron has its own cytoskeleton.What is its role? I am sure that
there is a great deal to be uncovered by future research,but it seems
that already a fair amount is known. In particular,microtubules in neurons
can be very long indeed,in comparison with their diameter (which is only
about 25-30nm) and can reach lengths of millimetres or more. Moreover,they
can grow or shrink,according to circumstances,and transport neurotransmitter
molecules. There are microtubules running along the lengths of axons and
dendrites. Although single microtubules do not seem to extend individually
to the entire length of an axon,they certainly form communicating networks
that do so,each microtubule communicating with the next ones by means of
the connecting MAPs referred to above. Microtubules seem to be responsible
for maintaining the strengths of synapses,and no doubt, for effecting alterations
of these strengths when the need arises. Moreover,they seem to organize the
growth of new nerve endings,guiding them towards their connections with other
nerve cells.
Since neurons do not divide after the brain is fully formed,there
is not a role of this particular kind for a centriole in a
neuron.Indeed,centrioles seem to be absent in the neuron's centrosome - which
is found close to the neuron's nucleus. Microtubules extend from there right
up to the vicinity of the vicinity of the presynaptic endings of the axon,and
also,in the other direction,into the dendrites and dendritic spines that
frequently form the postsynaptic end of a synaptic cleft. These spines
are subject to growth and degeneration, a process which seems to form an
important part of brain plasticity,whereby the overall interconnections in
the brain are undergoing continual subtle changes. There would seem to be
significant evidence that microtubules are indeed importantly involved in
the control of brain plasticity.
As an apparent curiosity,it may also be mentioned that in the presynaptic
endings of axons there are certain substances associated with microtubules
which are fascinating from the geometrical point of view,and which are important
in connection with the release of neurotransmitter chemicals. These substances
- called clathrins - are built from protein trimers known as clathrin
triskelions,which form three-pronged (polypeptide) structures. The clathrin
triskelions fit together to make beautiful mathematical configurations that
are identical in general organization to the carbon molecules
known as "fullerenes" (or "bucky balls") owing
to their similarity with the famous geodesic domes constructed by the American
architect Buckminster Fuller. Clathrins are much larger than fullerene
molecules,however,since an entire clathrin triskelion,a structure involving
several amino acids,takes the place of the fullerene's single carbon atom.
The particular clathrins that are concerned with the release of neurotransmitter
chemicals at synapses seem mainly to have the structure of a truncated
icosahedron - which is familiar as the polyhedron demonstrated in the modern
soccer ball!
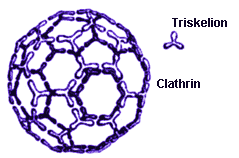 |
A clathrin molecule (similar in overall structure to a fullerene,but made of more complicated substructures - triskelion proteins rather than carbon atoms). The clathrin depicted resembles an ordinary soccer ball in structure. |
In the previous section,the important question was raised: what
is it that governs the variation in the strengths of synapses and
organizes the places where functioning synaptic connections are to be made?
We have been guided to a clear belief that it is the cytoskeleton
that must play a central role in this process. How does this help us
in our quest for a non-computational role for the mind? So far, all we seem
to have gained is an enormous potential increase in computing power over
and above what could have been achieved if the units were simply the neurons
alone.
Indeed,if tubulin dimers are the basic computational units,then
we must envisage the possibility of a potential computing power in the brain
that vastly exceeds that which has been contemplated in the AI literature.
Hans Moravec,in his book Mind Children (1988),assumed,on
the basis of a "neuron alone" model,that the human brain might in principle
conceivably achieve some 1014 basic operations per second,but
no more,where we consider that there might be some 1011 operational
neurons,each capable of sending about 103 signals per second.
If,on the other hand,we consider the tubulin dimer as the basic computational
unit,then we must bear in mind that there are some 104 dimers
per neuron,the elementary operations now being performed some 106
times faster,giving us a total of around 1024 operations
per second,as Moravec and others would strongly argue,there is no prospect
of the 1024 figure being achieved in the foreseeable
future.
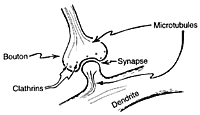 |
| Clathrins,like those above (and microtubule endings) inhabit the axon's synaptic boutons and seem to be involved in controlling the strength of the synaptic connection. |
Of course it could reasonably be claimed that the brain is operating
nowhere remotely close to the 100% microtubular efficiency that these figures
assume. Nevertheless, it is clear that the possibility of "microtubular
computing" puts a completely different perspective on some of the arguments
for imminent human-level artificial intelligence. Can we even trust
suggestions that the mental faculties of a nematode worm have already been
computationally achieved merely because its neural organization appears to
have been mapped and computationally simulated? As remarked earlier
the actual capabilities of an ant seem to outstrip
by far,anything that has been achieved by the standrad procedures of AI.One
might well wonder how much an ant gains from its enormous array of
nano-level "microtubular information processors",as opposed to what
it could do if it only had "neuron type switches". As for paramecium,there
is no case to answer.
Yet the arguments of Part 1 are making a stronger claim.I am
contending that the faculty of human understanding lies beyond any computational
scheme whatever. If it is microtubules that control the activity of the
brain,then there must be something within the action of microtubules that
is different from mere computation. I have argued that such non-computational
action must be the result of some reasonably large-scale quantum-coherent
phenomenon,coupled in some subtle way to
macroscopic behaviour,so that the system
is able to take advantage of whatever new physical processes must replace
the stop-gap R-procedure of present day physics.As a first step,we
must look for a genuine role for quantum coherence in cytoskeletal
activity.
Further Reading
|
| Chaos | Quantum | Logic | Cosmos | Conscious | Belief | Elect. | Art | Chem. | Maths |